PI System: Another Second Messenger
The phosphatidylinositol (PI) system is another second messenger system. PI is a minor component of membrane lipids. This molecule serves as a source of second messenger compounds. PI has three parts. See Figure 1.

Figure 1
The first part of PI consists of two fatty acids esterified to a glycerol. One of the fatty acids is the unsaturated fatty acid arachidonic acid (20:4), bound to carbon 2 of the glycerol. The other fatty acid is usually stearate (18:0). The combination of two fatty acids esterified to glycerol is called diacylglycerol, abbreviated DAG.
Another component of PI is a carbohydrate, phosphoinositol,which a phosphate diester binds to the third position of the glycerol. The inositol is usually phosphorylated at two positions.
Several types of signaling molecules are derived from PI. Hydrolysis of the glycerol‐phosphate linkage by a phospholipase leads to the signaling molecule trisphosphoinositol, abbreviated IP 3.
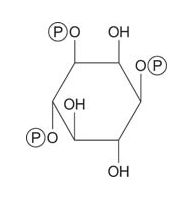
![]()
The remaining part of the phosphoinositol, 1,2‐diacylglycerol, is also a signaling molecule.
Finally, the arachidonate that arises from cleavage of phosphatidylinositol can serve as a precursor of prostaglandins.

Figure 2
Prostaglandins are mediators of the inflammatory response and are produced by the action of two activities of the enzyme prostaglandin synthase. The first activity is a cyclooxygenase activity, which adds two oxygen molecules the arachidonic acid. Secondly, the peroxide group from the first step is reduced to a hydroxyl group. The drug acetylsalicylic acid (aspirin) irreversibly inhibits the cyclooxygenase activity, while ibuprofen inhibits the reductase activity. Both drugs treat inflammation, pain, and fever because they inhibit prostaglandin synthesis. Prostaglandins are very unstable, so they tend to act locally (otherwise a sprained ankle would cause pain throughout the body).
IP 3 mobilizes Ca2+ from intra‐ or extracellular stores. The interior of a cell is kept very low in Ca2+ ions, at a concentration less than 10‐9 M., while the outside [Ca2+] is about 10‐3 M. This million‐fold concentration gradient is the result of cellular calcium‐dependent ATPase protein. Ca‐ATPase uses up to a third of the ATP synthesized by a cell to maintain the concentration gradient. The stores of Ca2+ available for use inside the cell are found primarily in the endoplasmic reticulum. A large store of Ca2+ exists in the mitochondrial matrix, but this seems to be a final “dumping ground”—in other words, calcium ions in the mitochondria don't come into the cytoplasm.
After Ca2+ comes into the cytoplasm, it binds to the mediator protein calmodulin. Calmodulin is a subunit of phosphorylase b kinase and a number of other enzymes. It binds Ca2+ with a K d of approximately 10 ‐6 M. When it does, calmodulin undergoes a conformational change; this conformational change activates phosphorylase kinase, which in turn leads to the activation of glycogen breakdown. Thus, an increase in intracellular [Ca2+] acts in the same manner as cyclic AMP. Phosphatases in the cell rapidly hydrolyze IP 3, which modulates the signal. The cytoplasmic Ca2+ is transported to the mitochondria and the cell returns to the resting state. The inositol is reincorporated into lipid and then re‐phosphorylated, ready to serve as a source of second messengers again.
Protein kinase C
After the phosphoinositol is released from the phosphatidylinositol, the remaining diacylglycerol is itself a second messenger, activating protein kinase C. The targets of protein kinase C are a number of intracellular proteins, including some that apparently target cell growth. Inappropriate stimulators of protein kinase C can be tumor promoters, compounds that stimulate the growth of cancer cells but do not cause genetic changes in the cell's DNA.