The Ferns and Their Allies
The seedless vascular plants are intermediate in their structural and reproductive adaptations between the more “primitive” bryophytes and the “advanced” seed plants. They often are called the amphibians of the plant world for although their sporophytes are well‐adapted to life on dry land, their gametophytes require a moist habitat to grow vegetatively and to reproduce sexually. The group includes the ferns and the “fern allies,” the latter a collection of plants whose relatives were the dominant plants in Paleozoic landscapes for 60 million or more years. Today, the members are a few remnant species reduced to an exceedingly minor role in the flora. The fossilized remains of the early vascular plants exhibit a variety of ways of coping with the terrestrial environment—only some of which were successful.
The relationships among and within the groups remain unclear for three major reasons: 1.) fossils form an incomplete record, but are the basis for many of the conclusions, 2.) data from molecular RNA sequencing of living species are incomplete, and 3.) opinions vary among botanists concerning how the available morphological and molecular data fit together in proposed phylogenies. The prevalent hypothesis concerning the placement of the groups on the large tree of life distinguishes two main lineages of vascular plants that diverge very early in the development of a land flora. One line includes the most primitive taxa and the lycophytes, the other the ferns, horsetails, and seed plants. The “which‐goes‐where” disagreements primarily concern extinct groups, but some extant ferns are problematical in their relationships also.
On one level the possession of vascular tissue—xylem and phloem—separates the ferns and their allies from the bryophytes and the lack of seeds from the gymnosperms and angiosperms. Other character‐istics they share in common are more varied and include:
- An alternation of a haploid gametophyte phase with a diploid sporophyte, a sporic meiosis. The gametophyte and sporophyte are nutritionally independent of one another.
- The sporophyte is the dominant, often branched, long‐lived phase (the leafy fern plant is the sporophyte, for example). Many are perennial and vegetative (asexual) reproduction is common.
- The gametophyte is smaller and either photosynthetic or saprophytic. Because the flagellated sperm need water in which to swim to the egg (like bryophyte sperm), the gametophyte is restricted in distribution by habitat. The plants are oogamous.
- Eggs are produced in archegonia, one per archegonium; sperm in antheridia, many per antheridium. The gametangia are multicellular with a protective coat of sterile cells and borne in nearby areas on one gametophyte or on separate ones.
- Haploid spores are produced by meiosis in sporangia and in some of the seedless vascular plants are of two different kinds: microspores and megaspores. The sporangia develop on specialized leaves called sporophylls. Some members of the group have strobili (singular strobilus), cones in which the sporophylls are clustered.
- A cell plate separates the new daughter nuclei during cell division.
- Cell walls are cutinized (unlike the bryophytes in which a cuticle is lacking).
- Xylem and phloem are well developed and transport water, minerals, and carbohydrates throughout the large sporophytes.
- Cellulose is the common wall material; a secondary wall of lignin strengthens the cells of most of the group.
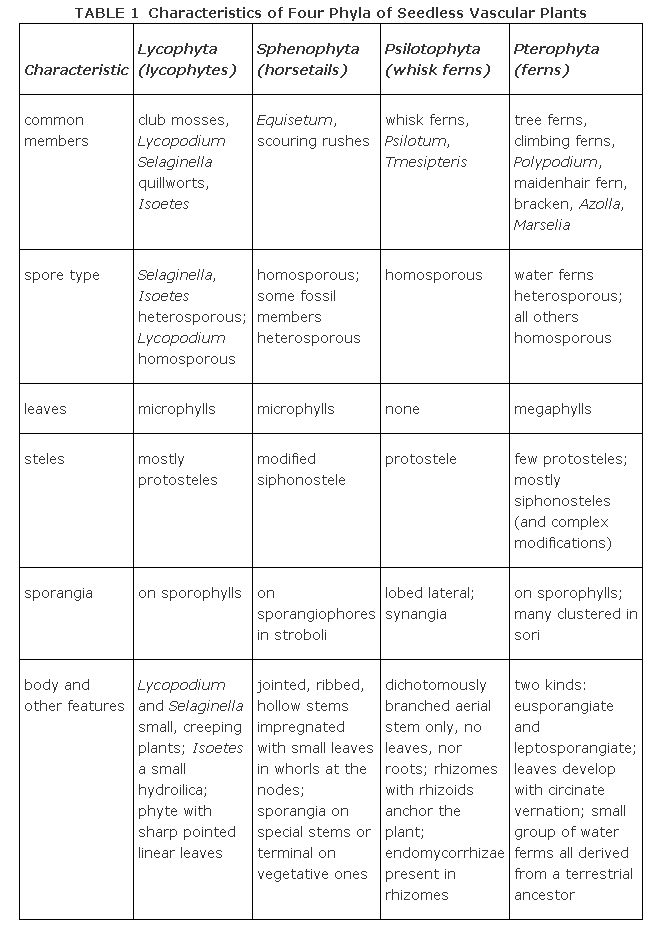
The living members of seedless vascular plants belong to four different phyla whose general characteristics are summarized in Table . Each group is described separately in the following pages.
|
|
|
|
|
|